4° Caso clinico: Disordini Temporomandibolari
 |
Article by: Gianni Frisardi
|
Introduzione
La letteratura epidemiologica indica che i disturbi temporomandibolari (TMDs) costituiscono una delle condizioni muscoloscheletriche croniche più comuni, con una prevalenza stimata tra il 5 e il 12% nella popolazione generale.[1] Studi basati su ampi campioni riportano che una percentuale significativa di soggetti riferisce dolore orofacciale, rumori articolari e limitazioni funzionali compatibili con TMD, senza che tali manifestazioni siano necessariamente associate a una patologia strutturale univocamente identificabile.
Dal punto di vista clinico, questa eterogeneità rappresenta una sfida rilevante. L’imaging riveste un ruolo centrale nella diagnosi delle patologie intra-articolari dell’ATM, ma non esaurisce la descrizione dello stato funzionale del sistema masticatorio e del network trigeminale. In molti pazienti, segni e sintomi sovrapponibili possono emergere in assenza di alterazioni morfologiche evidenti, suggerendo che la dimensione funzionale del sistema giochi un ruolo determinante.
Un aspetto critico riguarda l’interpretazione delle risposte neurofisiologiche trigeminali comunemente utilizzate nella pratica clinica. Riflessi quali il jaw jerk e il periodo silente sono noti per la loro sensibilità allo stato del sistema, al contesto sperimentale e al task motorio. La loro variabilità viene talvolta interpretata come rumore sperimentale o come indicatore indiretto di patologia, senza una chiara distinzione tra integrità strutturale e configurazione funzionale.
Questo capitolo utilizza un caso clinico con sintomatologia TMD-like come modello osservativo per descrivere tale distinzione. L’obiettivo non è validare una diagnosi specifica né proporre un approccio terapeutico, ma mostrare come, a fronte di una risposta evocata diretta stabile e riproducibile, le risposte riflesse trigeminali possano variare in modo significativo, riflettendo differenti stati funzionali del sistema neuromotorio.
Caso clinico

La paziente, di circa 40 anni, riferiva dolore orofacciale cronico, rumori articolari bilaterali dell’articolazione temporomandibolare e difficoltà masticatoria. Era già stata inquadrata come affetta da TMD secondo criteri clinici standard e sottoposta a trattamento conservativo. All’esame clinico si osservavano segni di disfunzione masticatoria e una malocclusione funzionale, senza la presenza di sintomi neurologici sistemici (vertigini, disturbi dell’equilibrio, acufeni).
In considerazione della persistenza dei sintomi e dell’assenza di elementi clinici suggestivi di patologia neurologica macroscopica, è stata eseguita una valutazione neurofisiologica trigeminale mirata, con l’obiettivo di distinguere
tra integrità strutturale della conduzione motoria e comportamento funzionale delle risposte riflesse.
Metodi neurofisiologici
La valutazione ha incluso:
- potenziali evocati motori bilaterali delle radici trigeminali (bRoot-MEPs) ottenuti mediante stimolazione transcranica;[2]
- registrazione del jaw jerk in posizione di riposo mandibolare;[3]
- registrazione del jaw jerk in condizioni di contatto dentario e attività muscolare moderata;
- valutazione del periodo silente dei muscoli masticatori evocato da percussione del mento durante serramento volontario.
Questa batteria è stata selezionata per confrontare una risposta evocata diretta, relativamente poco dipendente dal contesto, con risposte riflesse note per la loro modulabilità in funzione dello stato del sistema.
Risultati
Risposte evocate dirette (bRoot-MEPs)

Jaw jerk
Il jaw jerk registrato in posizione di riposo ha evidenziato asimmetrie in latenza e ampiezza tra i due lati. Tali asimmetrie risultavano accentuate quando il riflesso veniva evocato in condizioni di contatto dentario e attività muscolare, suggerendo una marcata dipendenza dal contesto e dall’input periferico.
Periodo silente
Il periodo silente mostrava caratteristiche compatibili con una modulazione funzionale del sistema, in particolare una riduzione o alterazione della fase di riattivazione motoneuronale successiva all’inibizione. Tali modificazioni non erano accompagnate da segni di alterazione della risposta evocata diretta.
In assenza di segnali suggestivi di alterazioni organiche della conduzione motoria trigeminale (bRoot-MEPs simmetriche), emergono caratteristiche compatibili con una modulazione funzionale del sistema:
- riduzione/alterazione della fase di riattivazione immediatamente successiva al periodo silente (rebound ridotto)
- profilo coerente con prevalenza di componenti inibitorie e con possibile ruolo di input nocicettivo e proprioceptivo nella modulazione fusimotoria
Meccanismi plausibili includono circuiti che coinvolgono afferenze nocicettive muscolari, subnucleus caudalis del V, interneuroni inibitori e modulazione di motoneuroni statici, con effetto sulla sensibilità dei fusi neuromuscolari.[4] [5] [6] [7] [8] [9]
Discussione
Il caso presentato evidenzia una dissociazione chiara tra due livelli di osservazione del sistema trigeminale. Da un lato, le risposte evocate dirette delle radici trigeminali risultano stabili, simmetriche e riproducibili, fornendo un riferimento di integrità strutturale della conduzione motoria. Dall’altro, le risposte riflesse — jaw jerk e periodo silente — mostrano una variabilità significativa, sensibile al contesto e allo stato funzionale del sistema.
Questa dissociazione non può essere interpretata come un artefatto sperimentale. Al contrario, riflette una proprietà intrinseca del controllo neuromotorio: le risposte riflesse non rappresentano output fissi, ma configurazioni emergenti di un sistema modulabile, influenzato da afferenze periferiche, tono muscolare, input nocicettivo e stato centrale.[10]
Nel contesto dei TMD-like, tale proprietà assume un significato clinico rilevante. La presenza di una risposta evocata diretta integra delimita il dominio strutturale del sistema, ma non consente, da sola, di dedurre lo stato funzionale espresso dalle risposte riflesse. Queste ultime possono variare in modo sostanziale pur in assenza di lesioni strutturali, rendendo problematica una lettura deterministica del segnale neurofisiologico.
Il capitolo non propone una soluzione a questo problema, ma lo rende osservabile. La coesistenza di stabilità evocata e variabilità riflessa rappresenta un punto di frizione tra misurazione neurofisiologica classica e interpretazione clinica, e costituisce un presupposto necessario per una lettura più articolata dei segnali neuromotori trigeminali nei quadri TMD-like.
Stabilità strutturale evocata e variabilità funzionale riflessa
La variabilità osservata nelle risposte riflesse trigeminali è coerente con quanto ampiamente descritto nella letteratura sulla neurofisiologia del controllo motorio umano. Studi sperimentali hanno mostrato che riflessi quali il jaw jerk e il periodo silente non rappresentano marcatori fissi di funzione, ma risposte dipendenti dallo stato del sistema neuromotorio, dal task e dal contesto di attivazione. In questo quadro, un’elevata variabilità non deve essere interpretata come rumore sperimentale o indicatore diretto di patologia, ma come espressione di un sistema adattivo e modulabile. La distinzione tra integrità delle vie neuromotorie e configurazione dell’output riflesso risulta quindi cruciale: una conduzione strutturalmente integra può coesistere con pattern riflessi variabili, che riflettono il controllo centrale in atto piuttosto che una disfunzione periferica.
La critica al peri-stimulus time histogram (PSTH) formulata da Türker e collaboratori [11] ha messo in evidenza come l’effetto sincronizzante dello stimolo possa contaminare la stima dei potenziali sinaptici indotti, rendendo necessaria l’introduzione di metodiche alternative e di procedure di normalizzazione analitica. Tuttavia, anche dopo la correzione della sincronizzazione, la stima del potenziale sinaptico netto rimane dipendente dal livello di attività pre-stimolo del motoneurone, riflettendo lo stato funzionale del sistema al momento della registrazione.
In questo contesto, risulta utile distinguere tra procedure di normalizzazione interne al segnale e riferimenti fisiologici strutturali. Le risposte evocate dirette delle radici trigeminali forniscono un’informazione relativamente indipendente dalla frequenza di scarica pre-stimolo e dal rumore sinaptico, delimitando il dominio di integrità della conduzione neuromotoria. Tale riferimento strutturale non sostituisce la stima dei potenziali sinaptici, ma consente di interpretare la variabilità delle risposte riflesse all’interno di un sistema la cui integrità di base è preservata.
Nel loro lavoro di sintesi sul controllo riflesso dei muscoli masticatori, Türker e collaboratori [12]hanno sottolineato come, negli esperimenti sull’uomo, non sia possibile stimolare un singolo sistema afferente in isolamento. Ogni condizione di stimolazione attiva simultaneamente più sistemi recettoriali, rendendo l’output riflesso intrinsecamente dipendente dal contesto e dallo stato funzionale del sistema. In questa prospettiva, la variabilità delle risposte riflesse non rappresenta un limite metodologico, ma una conseguenza strutturale dell’organizzazione del controllo neuromotorio umano. L’esigenza di standardizzare stimolo, registrazione e analisi nasce proprio dalla necessità di rendere confrontabili configurazioni funzionali diverse all’interno di un sistema che, sul piano della conduzione neuromotoria, rimane strutturalmente integro.
Nel contesto osservativo qui presentato, tale distinzione può essere resa operativa separando il dominio dell’integrità delle vie efferenti, valutato mediante risposte evocate dirette stabili e riproducibili, dal dominio dell’output riflesso, la cui configurazione rimane sensibile allo stato funzionale e al contesto di attivazione. Questa separazione consente di interpretare la variabilità delle risposte riflesse non come ambiguità diagnostica, ma come espressione di differenti stati del controllo centrale all’interno di un sistema strutturalmente preservato.
.
Un aspetto implicito, ma centrale, dell’impostazione proposta da Türker [13]riguarda il rapporto tra scala di osservazione e interpretazione funzionale. Se da un lato l’analisi a livello di singola unità motoria consente di ridurre gli errori introdotti dall’aggregazione del segnale e di stimare con maggiore precisione la modulazione sinaptica locale, dall’altro essa comporta inevitabilmente una perdita dell’osservazione del contorno funzionale globale. Gli stessi riflessi mandibolari, come sottolineato da Türker, non rappresentano l’attivazione di un singolo circuito, ma la risultante della co-attivazione simultanea di più sistemi recettoriali e vie afferenti, la cui integrazione emerge solo a una scala superiore. In questo senso, la discesa di scala migliora la risoluzione locale ma riduce la capacità di cogliere la configurazione complessiva dello stato neuromotorio. La variabilità dei riflessi può quindi essere letta non solo come espressione di modulazione sinaptica fine, ma anche come manifestazione emergente di un contorno funzionale che non è completamente accessibile attraverso l’osservazione microscopica isolata.
Alla luce del rumore sinaptico intrinseco, della variabilità neuronale riflessa e della natura reticolare dei network che condizionano l’output motoneuronale, emerge con chiarezza come il comportamento funzionale del sistema masticatorio non possa essere dedotto in modo univoco dalla sola configurazione morfologica. Le risposte riflesse non rappresentano la lettura diretta di una struttura, ma l’espressione emergente di stati funzionali dinamici, modulati dal contesto e dal livello di integrazione centrale.
Queste osservazioni rafforzano e rendono operativa la 'dissociazione tra forma e funzione', evidenziando il fallimento di un determinismo morfologico applicato a sistemi neuromotori complessi. In presenza di integrità strutturale delle vie efferenti, configurazioni funzionali profondamente diverse possono coesistere e manifestarsi attraverso pattern motori non sovrapponibili.
Il capitolo si colloca pertanto nel punto di frizione tra descrizione strutturale e osservazione funzionale, mostrando come la variabilità non costituisca un’anomalia da eliminare, ma una proprietà sistemica da riconoscere. È proprio questa dissociazione, resa evidente dall’analisi neurofisiologica, a indicare il limite interno della Scienza Normale e a preparare il terreno per un cambiamento di quadro concettuale che verrà affrontato nella sezione *Extraordinary Science*.
È opportuno distinguere in modo esplicito tra le categorie di malocclusione, trattamento ortognatico e disturbi temporomandibolari (TMDs), poiché esse non coincidono sul piano funzionale, pur condividendo spesso elementi morfologici sovrapponibili. La malocclusione, intesa in senso ortodontico, descrive una discrepanza geometrica tra arcate e basi scheletriche; la chirurgia ortognatica interviene su tale discrepanza modificando la configurazione strutturale del sistema. Tuttavia, né la presenza di una malocclusione né il suo trattamento chirurgico consentono di dedurre in modo deterministico il comportamento neuromotorio del sistema masticatorio.
Nei TMDs, il quadro clinico è ulteriormente complicato dal fatto che sintomi simili possono emergere in assenza di alterazioni morfologiche significative, mentre configurazioni strutturalmente corrette possono coesistere con pattern neuromotori instabili o asimmetrici. Il comportamento emergente del sistema masticatorio riflette quindi l’integrazione dinamica di molteplici network neuromotori e sensoriali, piuttosto che l’espressione diretta di una singola configurazione anatomica.
Questa dissociazione tra assetto morfologico e output funzionale evidenzia il limite di un approccio deterministico basato esclusivamente sulla forma. Il sistema masticatorio si comporta come un sistema complesso, nel quale stati funzionali differenti possono emergere da strutture simili e, viceversa, strutture differenti possono sostenere comportamenti neuromotori sovrapponibili. È in questo spazio di discrepanza tra forma e funzione che l’analisi neurofisiologica diventa necessaria per comprendere il reale stato operativo del sistema.-
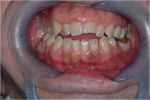 Figure 4: Paziente con marcata maloccusione dentaria. Dal punto di vista ortodontico, sarebbe improprio non considerare il paziente in una condizione di malocclusione. Secondo i canoni ortodontici tradizionali, si tratterebbe di un caso da trattare sia ortodonticamente sia chirurgicamente al fine di ripristinare una ipotetica “normocclusione”.
Figure 4: Paziente con marcata maloccusione dentaria. Dal punto di vista ortodontico, sarebbe improprio non considerare il paziente in una condizione di malocclusione. Secondo i canoni ortodontici tradizionali, si tratterebbe di un caso da trattare sia ortodonticamente sia chirurgicamente al fine di ripristinare una ipotetica “normocclusione”. -
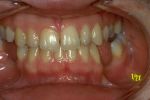 Figura 5: Paziente giunto all’osservazione presso il Dipartimento di Neuro-Gnatologia Funzionale, inviato da colleghi per riabilitazione protesica in mandibola edentula, considerata in condizioni di “normocclusione” dopo trattamento chirurgico ortognatico bimaxillare.
Figura 5: Paziente giunto all’osservazione presso il Dipartimento di Neuro-Gnatologia Funzionale, inviato da colleghi per riabilitazione protesica in mandibola edentula, considerata in condizioni di “normocclusione” dopo trattamento chirurgico ortognatico bimaxillare. -
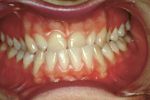 Figura 6: Situazione orale della paziente con sintomatologia TMD-like; si osserva crossbite anteriore.
Figura 6: Situazione orale della paziente con sintomatologia TMD-like; si osserva crossbite anteriore.


.
Ciò non mostra solo la variabilità neuromotoria e quanto le risposte riflesse trigeminali dipendano dallo stato del sistema neuromotorio, dal task e dal contesto di attivazione ma anticipa una concettualità concreta e cioè la 1️⃣ Dissociazione forma–funzione e fallimento del determinismo morfologico.
Se consideriamo i casi esposti in precenza (riportiamo soltanti dati più correlabili al tema in questione) possiamo notare come la funzione(rfilessi trigeminali) variano senza una correlazione morfologica vincolante. Il paziente
Conclusione
In presenza di integrità strutturale della conduzione motoria trigeminale, le risposte riflesse possono esprimere una variabilità ampia e dipendente dallo stato funzionale del sistema. Tale variabilità non deve essere interpretata come rumore, ma come espressione di configurazioni funzionali multiple del controllo neuromotorio.
La distinzione tra risposta evocata diretta e risposta riflessa emerge come elemento cruciale per comprendere la complessità dei quadri TMD-like e per delimitare correttamente il dominio di ciò che è strutturale rispetto a ciò che è funzionale.
- ↑ Ahmad M, Schiffman EL. Temporomandibular Joint Disorders and Orofacial Pain. Dent Clin North Am. 2016;60(1):105–124.
- ↑ Frisardi G. The use of transcranial stimulation in the fabrication of an occlusal splint. J Prosthet Dent. 1992;68(2):355–360.
- ↑ Cruccu G, Frisardi G, van Steenberghe D. Side asymmetry of the jaw jerk in human craniomandibular dysfunction. Arch Oral Biol. 1992;37(4):257–262.
- ↑ Ro JY, Capra NF. Physiological evidence for caudal brainstem projections of jaw muscle spindle afferents. Exp Brain Res 1999;128:425-434.
- ↑ Capra NF, Ro JY. Experimental muscle pain produces central modulation of proprioceptive signals arising from jaw muscle spindles. Pain 2000;86:151-162.
- ↑ Appelberg B, Hulliger M, Johansson H, Sojka P. Actions on g-motoneurones elicited by electrical stimulation of group III muscle afferent fibers in the hind limb of the cat. J Physiol. 1983;335:275-292.
- ↑ Macefield G, Hagbarth E, Gorman R, Gandevia SC, Burke D. Decline in spindle support to a-motoneurones during sustained voluntary contractions. J Physiol 1991;440:497-512.
- ↑ Mense S, Skeppar P. Discharge behavior of feline gamma-motoneurones following induction of an artificial myositis. Pain 1991;46:201-210.
- ↑ Pedersen J, Ljubisavljevic M, Bergenheim M, Johansson H. Alterations in information transmission in ensembles of primary muscle spindle afferents after muscle fatigue in heteronymous muscle. Neuroscience 1998;84:953-959.
- ↑ Kitagawa Y et al. Asymmetry in jaw-jerk reflex latency in craniomandibular dysfunction patients. J Oral Rehabil. 2000;27:902–910.
- ↑ Motor-unit firing frequency can be used for the estimation of synaptic potentials in human motoneurones. Türker KS, Cheng HB.J Neurosci Methods. 1994 Aug;53(2):225-34. doi: 10.1016/0165-0270(94)90181-3.PMID: 7823625
- ↑ Reflex control of human jaw muscles. Türker KS.Crit Rev Oral Biol Med. 2002;13(1):85-104. doi: 10.1177/154411130201300109.PMID: 12097240
- ↑ Estimating Exercise-Induced Changes in Human Neuronal Networks. Türker KS.Exerc Sport Sci Rev. 2021 Jul 1;49(3):147-156. doi: 10.1249/JES.0000000000000255.PMID: 33927162